Paul Grobstein
Department of Biology
Bryn Mawr College
Bryn Mawr, Pennsylvania 19010
(610) 526-5098 pgrobste@brynmawr.edu
published in The Encyclopedia of Human Behavior, Volume 4
(V.S. Ramachandran, editor), Academic Press, 1994 (pp 447-458)
copyright by Academic Press, Inc.
made available on Serendip
II. Behavior and the Nervous System: Input/output Boxes Within Input/Output Boxes
III. Intrinsic Variability in Input/Output Boxes at All Levels of Organization
IV. The Nature of Intrinsic Variability
V. The Analysis and Significance of Intrinsic Variability
VI. Conclusions
Information. An increasingly difficult and important term to define. Central to the concept is that there are aspects of reality which need to be characterized not in terms of matter or energy but rather in terms of the particular organized forms taken by one or the other or both. Similar information, for example, can be embodied as signals in motions of the air (sound waves), ink on a page (printed text), or a series of ones and zeros (on a hard disk in a computer).
Input/output device for information Any physical system whose primary function is to receive, transform, and emits signals (information, as opposed to matter or energy, see Information). Examples include thermostats, radios, computers, neurons, and the brain.
Levels of organization A term emphasizing the reality that virtually all interesting systems (the brain and behavior providing two examples) are made up of identifiable subsystems, which are in turn made up of identifiable subsystems, and so forth. As a general rule, the properties displayed at one level of organization reflect the properties of the subsystems at the next lower level of organization together with the particular way those subsystems are organized in relation to one another. The properties of the subsystems typically constrain but do not determine this organization (which reflects information over and above that inherent in the subsystems). For this reason, interesting system can productively be explored at all levels of organization, and need to be to be fully understood.
Membrane potential. The electric field which exists across the limiting membrane of a neuron (and many other cells). The value of the potential at any given time reflects the current and recent movement of charges across the membrane due both to intrinsic properties of the neuron and signals received from other cells. The value of the potential at any given time in turn determines the signals sent to other cells.
Nervous system The most sophisticated known input/output device for information. The term is a general one which, in animals with backbones (humans and frogs among them), includes the brain, spinal cord, and other related components. Other animals (including leeches) have nervous systems which, while somewhat differently organized, are fundamentally similar in terms of their basic function.
I. The problem of variability and its significance
A fundamental question about behavior, whether explored informally as all humans do all the time or formally as done under experimental conditions by scientists, is why a given organism at a given time under given environmental conditions behaves in a particular way. For many investigators, "understanding" behavior is equated with being able to predict for a given subject what behavior will occur at a given time. For others, the ability to study particular components of behavior depends on being able to reliably observe that behavior. In both cases, many investigators are predisposed to treat behavioral unpredictability as not only an inconvenience but, quite commonly, as a clear indication of the failure of the experimenter to control the circumstances with sufficient precision so that meaningful observations can be made.
This general perspective on variability has contributed importantly to a style of inquiry into behavior whose record of successes is both long and enviable. At the same time, it contributes to a dissociation, to the detriment of both, between the more formal and "scientific" study of behavior, and the common sense informal body of wisdom people are constantly creating in their day to day lives ("folk psychology"). Explanatory frameworks and models of behavior emerging from mindsets predisposed to dismiss variability tend, for obvious reasons, to have a deterministic character to them. These impact less than they might on folk psychology both because they appear somewhat threatening to human dignity, and because they fail to speak to a question central to informal understandings of behavior: they explain why a behavior occurs when it does, but not why it doesn't always occur. Conversely, the very real insights from the common sense body of wisdom about behavior, phrased as they are in terms of both puzzlement about its unpredictability and enjoyment of its playfulness, impact less than they might on more formal inquiries.
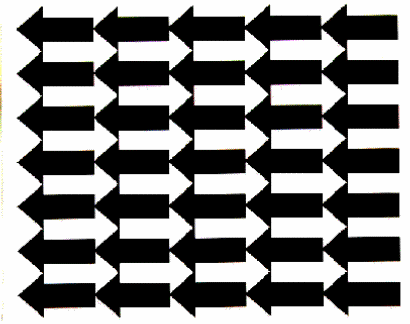
Mindsets, of course, influence the accumulation of data sets, and there is a corresponding bias againt the reporting of variable data sets in the scientific literature. This of course in turn influences mindsets, reinforcing those that dismiss variability as a meaningful and explorable phenomenon. Against this background, the existence of descriptions of behavioral variability in the biobehavioral literature is quite significant, and such descriptions do indeed exist. In the area of movement control, for example, there is a well-established body of phenomena under the rubric of "motor equivalence": under apparently identical task descriptions and circumstances, the particular movements made by even highly trained subjects vary for largely unknown reasons from occasion to occasion. Ambiguous figures, such as that shown in Figure 1, reveal a comparable largely mysterious variability in perceptual processing: while the particular figure seen can be influenced by prior suggestion, either form of the figure may be seen on successive presentations under apparently identical circumstances. Other examples where substantial variability has been documented include the processes of language acquisition in humans and song acquisition in birds. Both involve "babbling" phases during which largely unpredictable sound sequences are produced. More generally, the careful observational studies of ethologists have led some to suggest that the concept of a "fixed action pattern" should be replaced with that of a "modal action pattern" in recognition of the reality that even relatively stereotyped behavioral acts are not in fact identical when looked at more carefully.
In short, the scientific literature does in fact include fragmented supporting evidence for the Harvard Law of Animal Behavior. Despite this, there remains in many scientists' minds a general skepticism of the Law, and an associated reluctance to take it seriously. This is partly a methodological matter (how is one to study a mysterious variability?), and partly a pragmatic bet on the odds (if I work very hard to understand this variability it is just going to turn out to reflect my experimental inadequacies and disappear; besides, some imprecision and "noisiness" is inevitable in any real system). It also, however, reflects a deep-seated discomfort (not restricted to scientists) with the mysterious and uncontrollable, a combination of characteristics which smacks a bit too much of the supernatural. In the following sections are outlined a perspective which removes the taint of the mysterious and supernatural from the Harvard Law (without removing its core or its interest), providing as well both a methodological framework for further inquiry and a rationale for a pragmatic judgement that the phenomena are indeed worth exploration.
II. Behavior and the nervous system: Input/output boxes within input/output boxes.
Two general presumptions accepted to one degree or another by most neurobiologists underlie this discussion. The first is that behavior and the nervous system are in some sense the same thing: all aspects of behavior correspond in one way or another to phenomena of nervous system structure and function. The second general presumption is that an understanding of nervous system structure and function provides special advantages for better understanding behavior. An implication of the first general principle is that the Harvard Law ought to be understandable in terms of properties of the nervous system, and of the second is that knowing something of how the nervous system works should help to make sense of the Law. In this section is provided a description of some basic organizational features of nervous system structure and function relevant to these considerations.
To a good first approximation, the nervous system can be regarded as an input/output device for information which is made up of interconnected input/output information devices which are in turn themselves made up of input/ouput information devices. A convenient stopping point for what might, perhaps correctly, be regarded as an infinite regress is at the level of neurons, the cells which represent similar building blocks for all parts of the nervous system. Each neuron is itself an input/output device, receiving information (typically from other neurons) on receptor surfaces and transmitting information (typically to other neurons) from specialized effector regions. Two aspects of this picture are particularly important to appreciate, both at the level of the neuron and at the levels of each of the successive larger boxes of which they are a part. The first is that one can, with a fairly high degree of rigor, identify and independently monitor inputs and outputs. The second is that the input/output relationship of neurons is in general neither obvious nor stereotyped. It depends on intrinsic properties of the neuron itself, properties which can and do vary from neuron to neuron. That one can identify and independently monitor inputs and outputs, and in many cases control the former, provides the experimental ability to show the dependence of input/output relationships on intrinsic properties and, if one chooses, to characterize aspects of those properties, a point of substantial importance to what follows.
The same two principles hold for the nervous system as a whole, the highest level input/output box under consideration here (there is every reason to believe that a similar analysis could be usefully extended to still higher levels of organization, one in which individual organisms represent interconnected input/output boxes from which emerge, for example, properties of social organization). Like neurons, the nervous system has reasonably well-defined inputs (sensory neurons, the generally quite small percentage of the total neuronal population which consists of neurons whose receptor surfaces are outside the nervous system, rather than in contact with other neurons) and outputs (a second distinct set of neurons (including motoneurons), again generally a quite small percentage, whose effector regions are outside the nervous system, rather than in contact with other neurons). Here too, the input/output relations are neither obvious nor stereotyped (individual organisms behave differently from one another). They depend on intrinsic properties of the nervous system as a whole, a reality which can (as with neurons) be both verified and explored because of the ability to control inputs and monitor outputs.
Between the level of the nervous system as a whole, and that of neurons, there are a variety of levels of organization at which the description of interconnected input/output boxes each with significant intrinsic properties continues to hold. The neocortex, for example, has well defined input and output connections (restricted to other areas of the nervous system), and an internal organization which influences its input/output characteristics. So too does the spinal cord, which is interconnected by input and output pathways to other parts of the nervous system, and has, as well, associated sensory and motor neurons. These in turn are made up of smaller interconnected input/output boxes, each with characterizable and distinct intrinsic properties, such as different cortical regions and spinal cord segments associated respectively with forelimbs and hindlimbs. There is no unique way to describe the interconnected boxes within boxes which represent levels of organization between neurons and the nervous system as a whole, and it remains to be seen whether there is some best way. Regardless, the more general point holds. One can identify (perhaps inevitably with some degree of arbitrariness) at several levels of organization systems of related neurons with well-defined inputs and outputs, each having intrinsic organizations which are significant for determining their input/output characteristics.
III. Intrinsic variability in input/output boxes at all levels of organization
The discussion of basic features of nervous system organization in the previous section provides the basis for a useful rephrasing of the Harvard Law of Animal Behavior in terms which make it not only admissable but productive for scientific inquiry. What the Harvard Law in essence asserts is that there is some intrinsic variability in the intrinsic properties of the nervous system as a whole which influences its input/output relationships. To eliminate the taint of the supernatural, this property of the nervous system as a whole should be understandable in terms of the properties of the smaller boxes which make it up. To make the inquiry productive, it should be both explorable in terms of smaller boxes and meaningful for the understanding of the largest box (behavior). Central to all of this is the ability to specify rigorously what is meant by "intrinsic" variability, and to establish its existence.
The general point is both demonstrable and significant at the level of individual neurons. The output of a neuron is in general due to a time-varying membrane potential which most typically results in a time-varying pattern of action potentials and neurotransmitter release. Given that the inputs to a neuron are known, it is possible in principle (and, in many cases, in practice) to ask whether a time varying output pattern persists in the absence of any inputs (or at least of any time varying signal on the inputs). For many neurons, perhaps most, the answer is yes. Variations in membrane potential can persist in individual neurons totally isolated from any input pathways. Under normal circumstances, the outputs of these neurons reflect not only their inputs but also the membrane potential variations intrinsic to the neuron. To put the matter slightly differently, the Harvard Law of Animal Behavior holds for neurons. No matter how rigorously an experimenter ignorant of intrinsic variability controls the inputs to a neuron, its outputs for a given input can and, in many cases will, vary from trial to trial. While this may at first glance seem mysterious, the underlying cellular and molecular mechanisms are extensively studied and reasonably well understood. Suffice it here to say that within neurobiology it raises no eyebrows whatsoever to assert that the outputs of a given neuron reflect not only its inputs but also variations in signal processing characteristics which occur for reasons largely or entirely intrinsic to the neuron.
The ability to eliminate inputs and monitor outputs makes possible, at the level of the nervous system as a whole, a comparable demonstration and conclusion that one is dealing not with a simple input/output box, one in which a detailed knowledge of inputs suffices to predict outputs, but rather with an input/output box with intrinsic characteristics that vary with time. Figure 2A shows a continuous record of activity from the output nerves of the isolated nervous system of the leech, a preparation in which all input paths have been interrupted. It is clear that variations in output occur in the absence of any variations in input (and, indeed, in the absence of most if not all input of any kind). Preparations of this kind can in principle provide answers to the question that inevitably occurs to most students studying the brain: would a brain in a dish continue to think? The answer, at least in the case of the leech, is clearly yes. While much of the output activity in the isolated leech nervous system has unknown behavioral significance, some periods of this activity are clearly identifiable by their patterns as corresponding to behaviorally meaningful behavior. The rhythmic activity following the arrow in Figure 2A, for example, is the output pattern corresponding to swimming movements. Though comparablely controlled observations have not, for obvious reasons, been done with human brains, there is every neurobiological reason to believe that they too would exhibit variable and behaviorally meaningful output patterns despite isolation from inputs. In addition, "perceptual isolation" studies in humans clearly show that variations in brain states not only persist but may in fact be enhanced (yielding, for example, hallucinations) by reducing variations in input signals. That people frequently seek out quiet spots in which to think is unlikely to be a coincidence.
Figure 2B shows that the intrinsic variability present in the leech nervous system is relevant for understanding the Harvard Law of Animal Behavior. A cutaneous stimulus delivered to an intact leech sometimes, but not always, triggers swimming behavior. Such input/output variability might, in principle, be attributed to slight variations in the cutaneous stimulus itself, or to differences in other sensory inputs at the time the stimulus is delivered. In fact, as shown in Figure 2B, input/output variablity persists in the isolated nervous system. Identical electrical stimuli delivered to a peripheral nerve sometimes, but not always, triggers swimming. Notice also that, even when the stimulus does trigger swimming, there is substantial variation from trial to trial in the latency, swim robustness, and motor activity preceding the swimming episode. In short, the Havard Law of Animal Behavior holds for the nervous system as a whole.
Comparable observations have shown that intrinsic variability may also be a characteristic of the intermediate size boxes of which the nervous system as a whole is made. The significance of this will be discussed further below. Here it is only necessary to stress that neither the intrinsic variablity of the nervous system as a whole, nor that in any of the various intermediate boxes which make it up, require any appeal to the supernatural. Given intrinsic variability in neurons, there is every reason to expect its existence in the assemblies of neurons which make up both intermediate size boxes and the nervous system as a whole. In short, the failure of an experimenter to be able to predict the behavior of a given organism at a given time under particular environmental circumstances does not necessarily indicate either a failure of the experimenter to adequately control the circumstances nor an external deus ex machina. It may simply indicate a phenomenon which can be rigorously documented, the existence of intrinsic variation, and which can, as discussed further below, be taken as a legitimate subject of study in its own right.
IV. The nature of intrinsic variability
In the following, it will be useful to focus on the "nature" of variability, the particular form variability takes in a given situation, rather than on its causes. However, one subtlety in the definition of intrinsic variablity where issues of nature and cause are unavoidably intertwined needs to be dealt with first. As is evident from the various phenemona of learning and memory, the state of the nervous system at a given time depends not only on its inputs at that time but on the history of its past inputs as well. Such dependence can certainly contribute to difficulties in predicting the response of a given organism to a given input at a given time, but it is in principle at least a correctable problem (by due attention to controlling prior inputs, and/or to invariant correlations between prior and present events) and not the one of primary concern here. "Intrinsic variability" is intended to refer to changes in the state of the nervous system which occur largely or entirely for reasons independent of input signals at both present and prior times.
With this understanding, intrinsic variability can be conceived to take one of three forms: orderly, deterministic but ill-mannered, and probabilistic. Circadian rhythms, and other periodic phenomena known to be present in the nervous, neuroendocrine, and neuromuscular systems, provide clear examples of the first kind of intrinsic variability. Widespread, coordinated changes in the state of the nervous system on a roughly twenty-four hour cycle can persist for long periods of time in the absence of any driving variations in the input pathways of the nervous system, and there is every reason to think would occur in the absence of any experience at all with daily variations in input. They can occur as well in isolated parts of the nervous system, of various sizes down to the level of individual neurons. Periodic heartbeats are similarly largely independent of driving input at any point in time, and characterize the whole heart, as well as parts down to the level of single cells. Though not directly relevant to the concerns of this article, it is worth noting that, in both cases, the periodic intrinsic variability at the highest level of organization is not explained entirely by that at the lowest but depends as well on the interactions among those elements. Isolated heart cells may beat at frequencies quite different from each other and from that of the heart as a whole, which reflects not only the intrinsic properties of the individual heart cells but the interactions among them as well. Noteworthy too is that both heartbeat and daily activity cycles can indeed be affected by input signals. Orderly ntrinsic variability, indeed intrinsic variability in general, is a contributor to behavior, not an explanation of it.
There is a useful, if somewhat fuzzy, border between "orderly" and "deterministic but ill-mannered." Periodic intrinsic variability is well-studied at cellular and molecular levels, and raises no spector of the supernatural whatsoever. It also provides an equally safe route into the terrain of the "deterministic but ill-mannered". Any time varying signal, periodic or aperiodic, can be approximated as closely as one wants by the sum of a sufficiently large number of periodic signals (sine waves, for example) of different wave-lengths, amplitudes. and phase relations to one another. In particular, by summing the signal from a sufficient number of different elements, one can generate a signal (corresponding, for example, to the internal state of the nervous system) which is equivalent to the output of a random number generator. The resulting behavior would be deterministic, in the sense that it could be predicted if one had a knowledge of the underlying periodic elements. It can nonetheless be very "ill-mannered" in the sense that an outside observer, lacking a knowledge of the mathematics necessary to analyze the observed output, would see the behavior as highly or completely unpredictable. The category of "deterministic but ill-mannered" includes as well the various phemonena increasingly and popularly known as "chaotic." It is now well recognized that the behavior of even relatively simple systems involving small numbers of interacting non-linear elements (such as neurons) can be highly unpredictable. Such systems are unpredictable but determinate, in the sense that the time evolution of their activity can be calculated algorithmically given known starting conditions. The observed activity pattern itself, however, may, like that of summed periodic elements, exhibit little or no discernable order. An additional noteworthy feature of chaotic systems is a "strong dependence on initial conditions". Very small changes in the starting conditions can lead to very large differences in subsequent behavior. What this means is that chaotic systems, while determinate in principal, may not be so in practice. An observer is inevitably limited to a particular level of precision in the measurement of starting conditions, and the range within which the actual starting condition falls may well be early enough to yield a range of enitrely different outcomes.
In popular parlance, events are either predictable or unpredictable, with the former corresponding to "orderly" and the latter to "random" or "probabilistic." The recognition of the category of "deterministic but ill-mannered" is a generally important intellectual development which, however, seriously muddies this neat dichotomy. "Orderly", as already described, grades into "deterministic but ill-mannered," and there is some tendency to equate the the second with "unpredictable" in general. In fact, there is an important conceptual distinction between "deterministic but ill-mannered" and random or probabilistic. The latter terms, at least in their strongest sense, refer to systems in which it is presumed that the unpredictability is not a matter of practice but instead one of principal. Quantum mechanics provides the most explicit modern conceptualization of the intuition that such in principal unpredictable systems exist; coin flipping or dice-throwing are vernacular metaphors for the same intution that some phenomena have a truly indeterminate character. It is not at all clear at the moment whether there exists an operational criterion by which an observer can distinguish between variabilities which are "deterministic but ill-mannered" as opposed to probabilistic. Nonetheless, the distinction is conceptually clean, and has at least one quite important general implication, as discussed further below.
Orderly intrinsic variability is a well documented aspect of nervous system function. So too is less orderly intrinsic variability in neuronal activity, falling into the general area of the second two classes mentioned. There exists an older literature describing "noise" in both single neuron activity and in such monitors of global activity as the EEG (such observations were not always done under conditions adequate to rigorously demonstrate that the activity was intrinsic, but such is very much consistent with the trend of the evidence). More recently, evidence of chaotic activity has been obtained in a variety of systems, including invertebrates as well as the olfactory bulb and neocortex of mammals. While there are varying degress of uncertainty (both practical and conceptual) in these observations, there is no question but that a substantial degree of unpredictability characterizes the physiological activity of many parts of all nervous systems under conditions of substantially controlled input activity.
V. The analysis and significance of intrinsic variability
Preceding sections have provided neurobiological evidence in support of the notion that the Harvard Law of Animal Behavior may reflect actual variability in nervous system function, rather than supernatural forces or the failure of investigators to adequately control experimental circumstances. What remains for discussion is whether this understanding yields productive new lines of investigation. Two questions are at stake. One is whether there are ways to usefully explore observed variability, and the other is whether variability has any functional significance.
V.1. Methodology: a case study in frog and leech
Figure 3A provides a concrete illustration of the Harvard Law of Animal Behavior, drawn from studies of frog prey orienting behavior. In each of a series of trials, a live prey item was confined in a cup at a fixed angle and distance from a given frog. The frog oriented accurately on all trials, in the sense that a line corresponding to the midsaggital plane of the head passes, when extended outward, through the cup containing the prey item. The actual position and head angle, however, varies substantially from trial to trial, as shown in the figure. Most people's presumption is that such variability relates to a trial to trial variability in the input signals, an obvious possibility being a variation in those related to prey location (which can vary somewhat within the cup) or in the frog's posture at the time the stimulus was presented. The variability is, however, neither decreased by decreasing the size of the cup (Figure 3B), nor increased by causing the frog to adopt a wider than normal range of initial postures (Figure 3C). These findings do not preclude some more elaborate explanation of the observed variance in terms of variations in input signals. They do, however, provide the kind of evidence (equally attainable in humans) which would support further consideration of the likely existence of intrinsic variability, and motivate additional experiments (of the kind discussed in previous sections where possible) to try and rigorously establish its existence.
Even given such a demonstration, many people might discount the significance of the observed variability, on the grounds that the nervous system is a complex physical device and that a certain imprecision in the function of any such device is inevitable. That the observed variation has an organized character (the frog is always pointing at the prey item) rather than being random provides some argument against this interpretation but an even stronger case can be made (here and in other situations). The observed variability is not constant, but instead varies depending on the circumstances. In a more complex visual environment (Figure 3D), the variability is less. This clearly indicates that the variability evident in Figure 3A is not simply that inherent in the limits of precision of a physical device, but rather an independently variable property of the physical device.
The same is true for the rigorously verified intrinsic variability described in an earlier section for the leech. Figure 2C illustrates the stimulus/response curve for the isolated nervous system, showing that over a substantial range of stimulus voltages, the response in terms of the presence or absence of swimming is unpredictable (response percentages neither zero nor 100%). The behavior is much more predictable when the head ganglion is removed from the nervous system, as show in Figure 2D. For each stimulus voltage the preparation reliably swims or fails to swim. Here too the intrinsic variability is not an expression of a constant inevitable imprecision but rather a characteristic which can be reduced under certain circumstances.
The findings in the leech further suggest that different regions of the nervous system (different intermediate size boxes) may play different roles in intrinsic variability, with some areas increasing it and other areas decreasing it (removal of the tail ganglion in the leech tends to increase observed variability). Interestingly, the history of neurobiological research, viewed in this light, suggests a similar conclusion. Experimental neuroscientists, interested in stable and predictable preparations in which to explore the details of particular pathways, long ago discovered that removal of the more rostral parts of the nervous system yields more reliable input/output relations in the spinal cord. While it is possible that this increased stability results from elimination of sensory input paths associated with the more rostral parts of the nervous system, it is equally possible that the observed reduction in variability would also be observed in a totally isolated nervous system. This possibility seems worth serious attention, and may relate to the experience of skilled athletes, whose performance at times seems to depend on blocking out not only input signals but their own thinking as well.
The general problem of how the nervous system is organized so as to potentially and somewhat unpredictably associate a given input with any of a number of outputs similar in overall objective but differing substantially in detail is a wide open area for new investigation. Related to this are a series of open questions about the nature of the variability itself (some form of orderly but ill-mannered, or statistical?) and how it is created (molecular noise or something at a higher level of organization?). Clearly, an inquiry proceeding from a presumption of the likely existence of intrinsic variability is not only both achievable and opens new lines of scientific investigation, but can usefully bridge between the later and folk psychology as well.
.
V.2. Significance
The physicist Erwin Schrödinger, in his classic What is Life?, remarked fifty years ago on the remarkable stability and predictability of biological systems, noteworthy not only in comparison to the thermal noise of all physical systems but also because the biological systems are, of course, made up of noisy physical elements. From Schrödinger's perspective the problem of biological systems was one of how to combine physical elements so that their inevitable noise was reduced in the combination. The discussion to this point suggests that aspects of nervous system organization may well have evolved with exactly the opposite tendency: to enhance some kinds of noisiness or variability. There is, in fact, some reason to believe this is true of biological systems in general. If intrinsic variability is not actually the residual noise of imperfect machinery, the question to be treated at this point is what functions might it serve?
There is, across the sciences generally, an increasing interest in, and inquiry into the properties of "complex systems." Associated with this is the emergence of new and useful metaphors to try and imagine how properties like those displayed by the brain can emerge from simpler elements. "Complex systems" in general display substantial form and organization in the absence of anything corresponding to either an external planner or an internal blueprint; their properties instead emerge simply from the interactions of large numbers of elements. In this sense, neither the "machine" explanatory metaphor of the nineteenth century nor the "computer" metaphor of the later part of the twentieth is appropriate.
A more useful metaphor for "complex systems", suggested by some workers in the area, is that of a sandpile, as it might emerge from a continuous flow of sand dropping onto an elevated dish. The sandpile will grow until it reaches a particular size and shape. Despite the absence of any planner for the sandpile, either external or internal, its subsequent size and shape will persist more or less unchanged, as comparable amounts of sand enter and leave the sandpile by falling off the edge of the dish. The sandpile is an "open" and "self-organizing" system. Its form depends on a continuous flow of matter and energy through it, and emerges entirely from the interaction of large numbers of elements. Such systems in general display both homeostatic and adaptive properties. If the pile is disturbed by flattening it, it will, over time, return to its original form. If the dish is made larger, the sandpile will, over time, modify its shape and form to make use of the additional space. Most importantly, in the present context, such open, self-organizing systems are in general "noisy." While the overall size and shape of the sandpile is constant, there are continous and substantially unpredictable small changes in its form: at one point the addition of a few grains of sand results in a comparable number of grains leaving the pile, at another point the same few grains added triggers a major avalanche. Despite the appearance of relatively constant form, the sandpile is in fact constantly in a state of flux, indicated by its noisiness, and it is in fact precisely because of this flux that the sandpile displays both homeostatic and adaptive properties.
The sandpile is an instance of a "determinate but ill-mannered system", one in which substantial unpredictability in practice emerges as a property of the quite determinate interaction of quite determinate elements. It is a particularly simple example of such a system, having only one relatively stable form. More interesting, but not enormously more elaborate, systems can have many relatively stable forms, and shift among them relatively unpredictably (imagine an appropriately sized hole in the center of the dish, so that every once in a while, the pile collapses in the center, changing from a mountain into a volcano). Nonetheless, the sandpile serves to illustrate an important general point: that variability is not only not inconsistent with either homeostasis or adaptability but in fact reflects precisely those phenomena (large numbers of interacting elements in an open system) which gives complex systems their adaptive and homeostatic characteristics. Phenomena of both perception and movement control are currently being usefully explored from this perspective. Object recognition, for example, may be best thought of as a variability-added search for a stable configuration of a large number of interacting elements (two such roughly equivalent stable points corresponding to the skull and woman with mirror of Figure 1).
There are, however, additional and more direct possible significances of "noisiness" which should also be born in mind. Game theory illustrates one such. The success of an organism at any given task is frequently dependent on the playing out of interactions with other organisms, interactions in which the behavior of each influences the behavior of the other. If the nature of the task is such that greater success of one organism necessarily implies lesser success of the other, then there exists an optimal strategy for the "game", a well-defined set of rules which will assure the maximal achievable score, assuming both participants behave "rationally." In many cases (poker providing one example), it can be proven that this optimal strategy includes deliberate randomization of behavior. Variability in the frog's prey orienting movements may, for example, make it more difficult for the prey to successfully escape.
Variability can also be used to achieve an objective in the absence of existing information about exactly how to achieve that objective. Here the chemotactic behavior of many protozoans is instructive. In the absence of a chemical gradient of an attractive substance, these organisms move continuously with intermittent and apparently random changes of direction. The frequency of direction changes is, however, a function of the concentration of attractive substances, decreasing with higher concentrations. Because of this, an animal in a concentration gradient will reliably end up at the source, always, of course, by a different path. There is some evidence that frogs, once they acquire information about target location, may have a related capability to use their variability to create novel ways of reaching that target when more straightforward methods are unavailable. Indeed such a "creative" use of available variability may be a quite general property of movement control systems which display "motor equivalence", and perhaps contribute to explaining how it comes to be that years on a playground basketball court can yield performances never before seen.
The discovery, exploration, and creation of novelty has, of course, significance beyond the restricted domain of movement control, and in this wider terrain too variability plays obvious roles. "Noise" is an important element in artificial intelligence systems, where it is used to prevent premature settling on a less good solution to a task so as to ensure the search continues until better ones are found. Most humans have similar experience with needing to "get out of a rut" in their own day to day lives. More generally, substantially unpredictable variability must necessarily underlie all genuinely creative processes, since by definition they represent ways of dealing with phenomena for which the underlying rules are unknown. Dreams provide a familiar archetype of this kind of variability: while the elements in dreams (blue, three horns, snake, woman's head) are frequently familiar, the particular ways they are put together (a blue three-horned woman's head on a snake's body) frequently display a substantially unpredictable variability. The experiences one has with such things represent useful (and used) explorations of the novel, and the unpredictable combinations themselves sometimes emerge as quite significant insights (the "Eureka!" phenomenon).. .
The preceding is intended to establish that variability is by no means the residium left over when everything else is understood, but may instead in many cases be part of the very essence of what is to be explained, since many aspects of behavior relate more or less directly to problem solving, exploration, and creation. It is not intended to be a complete catalogue of the possible significances of intrinsic variability, and the issue of whether one is dealing with "determinate but ill-mannered" as opposed to probabilistic has deliberately been left vague for the cases discussed. If there is in fact a real distinction between a sufficiently chaotic process and a random number generator, it is probably not a distinction which matters for the situations described (though this is in fact a matter worth exploration). There is, however, one additional phenomenon worth mentioning in a discussion of variability where the distinction is in fact critical.
The existence and nature of "free will" is a major unsettled issue in western philosophy, one for which the Havard Law of Animal Behavior is relevant. Roughly speaking, the question is whether the output of a given organism is under its own control, as opposed to being fully determined by in-principle-knowable causal factors. In neurobiological terms, one component of the question can usefully be rephrased in terms of whether complete information about the genome of an organism, together with similarly complete information about all input signals which the organism has received during its lifetime, would suffice to make the next output predictable. The discussion of intrinsic variability given to this point implies that the answer to this question is no: even identical twins reared under identical circumstances would behave differently if a sufficiently chaotic intrinsic variability influenced their output, either directly or via the longer lasting effects of intrinsic experiences associated with dreaming or thinking. And they would certainly behave differently if the relevant intrinsic variability was truly probabilistic. This frees one to entertain the possibility that behavior is not fully determined. Finding oneself behaving unpredictably does not, however, fully satisfy the notion of "free will." The remaining question is that of control. Does an individual organism in fact have any control over its output?
A reconsideration of Figure 1 suggests that some organisms, at least, can in fact exert meaningful control over their own output. As earlier described, the figure can, somewhat unpredictably, be seen as arrows pointing either to the right or to the left . Most individuals, staring at the figure for a sufficiently long time, can in fact see it at one time as one and at another time as the other. It is also possible for humans (at least) to withhold output (in response, for example, to the question "what do you see?") until the picture is seen as one or the other of the two images. One is, in short, capable of monitoring the consequences of the intrinsic variability of one's own nervous system, and withholding an output until that variability yields something with which one is satisfied. Individual organisms, including individual humans, are both made up of and are a part of complex causal webs of interacting elements. That they are fully under their own control is clearly not the case. The present discussion, however, suggests that the combination of intrinsic variability and a monitoring/editing function may indeed endow at least those organisms which possess the combination with a quite satisfying degree of "free will."
VI. Conclusions
The Harvard Law of Animal Behavior clearly warrants more serious attention than it usually receives. A reasonable skepticism about observed behavioral variability, due to the possible existence of unknown but identifiable uncontrolled variables, is both reasonable and demonstrably productive as a research strategy, insofar as it results in the identification of new causal influences. At the same time, the phenomena of intrinsic variability discussed here provide viable alternative explanations of behavioral variability, potential explanations which are at least as likely, rigorously explorable, and significant as an unknown uncontrolled but potentially controllable variable.
Two more general implications of this conclusion are worth making explicit. The first is that the equation between "understanding behavior" and "being able to predict the behavior that a given organism will display at a given time", no matter how obvious it might seem nor how useful in particular inquiries, is fundamentally flawed as a general prospectus for the behavioral and biobehavioral sciences. Some degree of unpredictability is, as suspected by the common sense view, a fundamental aspect of behavior. This raises some interesting questions about the meaning of "understanding" in the context of the scientific study of behavior, but other disciplines, notably physics, have managed to incorporate "uncertainty" into their prospectuses, and there is no reason to believe that the scientific study of behavior can not successfully incorporate a "biobehavioral uncertainty principle" as well. Acknowledgement of such a principal would, in any case, contribute to a rapprochment between the common sense and scientific explorations of behavior, reducing the perceived threat to human dignity associated with a presumption that behavior (including that of humans) can in principlebe fully predicted, while at the same time making clearer the successes inherent in being able to say why an organism behaves the way it does when it does. Intrinsic variability not only removes the spectre of absoute predictability, but may, as discussed, provide a basis for admitting more fully into scientific discourse the concept of free will.
The second, important general implication is that intrinsic variability is not necessarily "noise", in the sense of being an inevitable byproduct of any real system, but instead is, in many cases at least, an essential ingredient of successful behavior. Scientists tend to focus on the "adaptiveness" of behavior, on the underlying organization which yields an "optimal" behavior in a given context. What game theory establishes is that, in many contexts at least, "optimal" is not a particular behavior, but rather variable behavior. Even more importantly, however, what is forgotten in the emphasis on "adaptiveness" and "optimality" is that much of behavior is fundamentally exploratory. Even when the task is known, as in the case of frog orienting, the underlying neuronal organization seems to be organized so as to explore various ways of achieving that task. To put the matter differently, "play" (and its associated behavioral variability) is not purely entertainment or a luxury to be given up when things get serious. It is itself a highly adaptive mechanism for dealing with the reality that the context for behavior is always largely unknown. Given that the adaptiveness of behavior itself derives from an evolutionary process in which variability and play are absolutely essential (and that no more successful mechanism for creating adapted systems is known), this should not be surprising. Scientists have, however, tended to focus more on the selection process than on the underlying variability without which it cannot act. In any case, recognition of intrinsic variability in the nervous system provides a second basis for rapprochment between the scientific and common sense perspectives on behavior: playfulness is indeed not only to be enjoyed but to be accorded high value for its fundamental role in the success of all organisms, including humans.
BIBLIOGRAPHY
Abbs, J.H. and Cole, K.J. (1987) Neural mechanisms of motor equivalence and goal achievement. In: "Higher Brain Function" (S.P. Wise, ed.), Wiley, New York, pp 15-44.
Bak, P. and Chen, K. (1991) Self-organized criticality. Scientific America, January, pp 46-53.
Barlow, G. W. (1977) Modal action patterns. In: "How animals communicate" (T.A. Sebeok, ed.) Indiana University Press, Bloomington, Indianna, pp 98-133.
Berkinblitt, M.B., Feldman, A.G., and Fukson, O.I. (1986) Adaptability of innate motor patterns and motor control mechanisms. Behav. Brain Sci. 9: 535-638.
Grobstein, P. (1988) From the head to the heart: some thoughts on similarities between brain function and morphogenesis, and on their significance for research methodology and biological theory. Experientia 44: 960-971.
Grobstein, P. (1992) Directed movement in the frog: motor choice, spatial representation, free will? In "Neurobiology of Motor Programme Selection" (J. Kien, C.R. McCrohan, and W. Winlow, eds.), Pergamon Press, New York, pp 250-279.
Kauffman, S.L. (1993) "The Origins of Order", Oxford University Press, New York.
Linsker, R. (1990) Perceptual neural organization: some approaches based on network models and information theory. Ann. Rev. Neurosci. 13: 257-281.
Llinás, R. (1990) Intrinsic electrical properties of nerve cells and their role in network oscillation. Cold Spring Harbor Symp. Quant. Biol. 55: 933-938.
Pittendrigh, C.S. (1993) Temporal organization: reflections of a Darwinian clock watcher. Ann. Rev. Physiol. 15: 17-54.
Poundstone, W. (1993) "Prisoner's Dilemna" Doubleday, Anchor Books, New York
Ruelle, D. (1991) "Chance and Chaos" Princeton University Press, Princeton, New Jersey.
Shepard. R.N. "Mind Sights" (1990) W.H. Freeman and Co., New York.
Skarda, C.A. and Freeman, W.J. (1987) How brains make chaos in order to make sense of the world. Behav. Brain Sci. 10: 161-195.
FIGURE LEGENDS
Figure 1. An ambiguous figure. Created by the author based on a figure by Roger Shepard in Mind Sights (W.H. Freeman and Company, 1990). The figure, sometimes seen as a series of arrows pointing to the left and other times as a series of arrows pointing to the right, provides a striking example of the importance of an appreciation of intrinsic variability for the understanding of behavior (see text).
Figure 2. Intrinsic variability in the leech nervous system. A. A record of continuous electrical activity from one peripheral nerve of an isolated leech nervous system. The observed activity varies with time despite the absence of variations in input, and includes periods of activity known to be associated with well defined output behaviors such as swimming (the bursting activity beginning at the arrow). B. Variability in response to a fixed stimulus. Shown are five trials on each of which the same electrical stimulus (line below bottom trace) was applied to a nerve other than the one being recorded from. Notice that the same stimulus sometimes, but not always (B4) causes swimming. Even on trials when swimming does result, there is substantial variability in the pattern and duration of activity prior to the initiation of swimming , as well as in the swimming activity itself (B1,2,3,5). C. A stimulus response curve for the isolated leech nervous system. For each of a series of stimulus voltages (abscissa) the figure shows the percentage of trials (ordinate) using that voltage on which a swimming episode resulted. Notice that, over a wide voltage range, the same stimulus sometimes does and sometimes does not trigger a swim. D. A stimulus response curve for the isolated leech nervous system in the absence of the most head ganglia. Conventions as in C. Notice that there is now a sharp transition between stimulus voltages which reliably do and do not trigger swimming episodes. Unpublished studies by Peter Brodfuehrer and Paul Grobstein.
Figure 3. Behavioral variability in the frog. A. Base line behavior. On each of ten trials, the frog was positioned at the lower cross (long vertical line is body axis, short horizontal line the line between the frog's eyes) and presented with a live mealworm confined in a cup (short upper horizontal line.Thinner oblique lines show the position and angle of the frog's head after the first movement on each trial. Notice that both position and angle vary substantially. B. Reduction of stimulus cup size (shorter upper horizontal line). Notice that there is no corresponding reduction in the variability of final head position. C. Increase in variability of initial posture (broadening of line corresponding to body axis). Notice that there is no corresponding increase in the variability of final head position. D. A richer visual environment. Notice that, under this condition, the observed variability in final head position significantly decreases. Unpublished studies by Paul Grobstein, Jeff Oristaglio, and others.
{kind=link}